| HOME |

|
|
|
Modifications to the Cactus Cortex The changes that occurred in the shoot cortex during cactus evolution involved more modifications than just adding more water-storage capacity by adding more parenchyma cells. The evolutionary increase in width of cactus stems due to having an increasingly wider cortex affected the selection pressures acting on numerous aspects of cactus biology, including: 1) the need to translocate water and sugar within the cortex itself; 2) the ability to change volume without changing surface area, 3) the mechanical tissues needed to support the increased weight of stems, and 4) the number of shoot apical meristem cells needed to produce the numerous cells of the voluminous body.
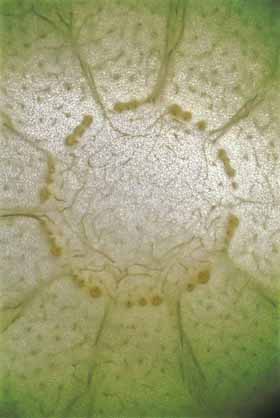
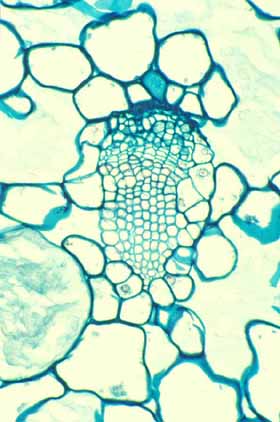
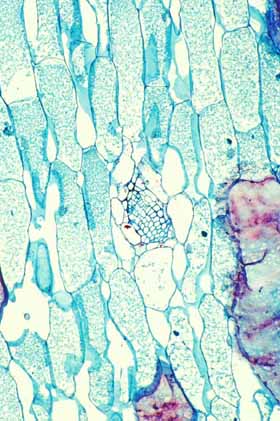
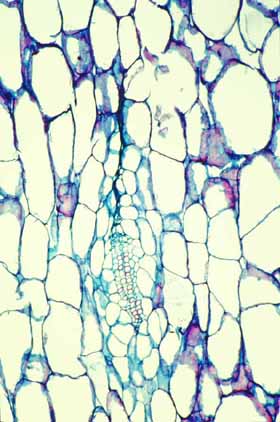
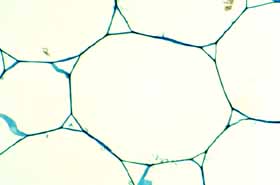
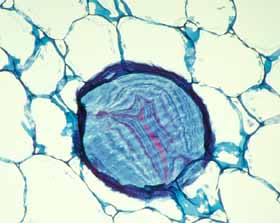
Cortical bundles: The translocation of water and sugar through the voluminous cortex One consequence of the increased cortex width is that the photosynthetic outer cortex is pushed much farther away from the vascular bundles of the stem (the cauline bundles) as the cortex differentiates. Most cacti still have a relatively narrow pith, so in broad, globose barrel cacti and those with wide columnar shoots, the epidermis and chlorenchyma are often several centimeters – and as much as 30 cm in Echinocactus – from the xylem and phloem of the shoot’s vascular bundles. Over such distances, diffusion of water is extremely slow, diffusion of glucose or sucrose is even slower. Even with an extremely thick covering of cuticle and wax, any epidermis will lose some water to dry desert air, and that water must be replaced rapidly if the epidermis and outer cortex tissues are to remain alive. Similarly, as the outer chlorophyll-bearing cortex cells produce sugars and starch, those carbohydrates must be moved away from the plant’s surface if they are to be of any use to the rest of the plant. Rapid translocation throughout the voluminous cortex of many cacti is made possible by the presence of a system of collateral cortical bundles. Cortical bundles branch off from stele bundles and from leaf/bud traces, then run outward, tangentially and upward through the cortex, branching extensively and vascularizing all regions of the cortex except the outermost layers of palisade cortex. Cortical bundles always end near or in the basal layers of the palisade cortex, they never extend to the hypodermis or epidermis. As with leaf venation, the most distal portions of cortical bundles are small, narrow bundles with few conducting cells, so that they look like leaf minor veins. In a large number of species, cortical bundles end in clusters of cells that resemble terminal tracheids in leaf veins: the cells are large and irregularly isodiametric but they have helical or reticulate secondary walls reinforcing their primary walls. These “terminal tracheid-like” cells usually occur in small clusters of about ten to twenty cells.
Because cactus cortex is long-lived, cortical bundles must also function for many years. In all species, cortical bundles produce secondary phloem, and in very old bundles at the base of very old shoots, the amount of collapsed secondary phloem can be many millimeters thick. Many species also produce secondary xylem in their cortical bundles, but typically it is only a small amount even in very old bundles in very old cortex. An unusual feature that needs more study is that in a few species, cortical bundle secondary xylem contains a matrix of xylary fibers, not xylary parenchyma.
Because cortex of most cacti is vascularized by cortical bundles, the epidermis and palisade chlorenchyma can be kept hydrated and healthy no matter how much water they lose to the dry desert air, as long as the plant itself has enough water of course. The presence of cortical bundles may be a key innovation in the evolution of cacti because it has permitted cactus cortex to become extremely broad as mentioned above. It is hard to over-emphasize the importance of the evolution of cortical bundles in cacti: whereas we animals vascularize any additional tissues we produce (such as tumors), plants do not do this automatically. In contrast, a plant’s ability to produce extra tissues is extremely limited by its inability to vascularize them. Consequently other stem-photosynthetic plants such as euphorbias, pachypodiums, stapeliads never become as broad as cacti do. It is certainly popular to point out the convergent evolution between euphorbias and cacti, but no euphorbia ever becomes as broad as a barrel cactus, or even as broad as many smaller cacti. The reason is simple: none of the non-cactus stem-photosynthetic succulents have cortical bundles in their stems. They are free to evolve to have a wider, more voluminous cortex only up to the point at which diffusion from the stele becomes too slow to keep the epidermis healthy. One of the broadest euphorbias, E. obesa, achieves its width by widening its stele: it has a broad pith that spreads its set of vascular bundles into a ring with a wide diameter, which is in turn surrounded by a cortex that is only moderately thick. But storing water in pith is more difficult than storing it in the cortex because a pith can absorb a large amount of water and swell only if the wood is flexible, otherwise either the wood would be torn apart or the pith would be prevented from expanding enough to absorb much water. Cortical bundles are present in almost all members of subfamily Cactoideae (only Blossfeldia liliputana lacks them) but are absent from subfamilies Pereskioideae and Opuntioideae.
Mauseth, J. D. 2004. The structure of photosynthetic, succulent stems in plants other than cacti. The International Journal of Plant Sciences 165: 1-9. Mauseth, J. D. 2004. Cacti and other succulents: stem anatomy of “other succulents” has little in common with that of cacti. Bradleya 22: 131-140. Sajeva, M., and J. D. Mauseth. 1991. Leaflike structure in the photosynthetic, succulent stems of cacti. Annals of Botany 68: 405 – 411. Mauseth, J. D., and M. Sajeva. 1992. Cortical bundles in the persistent, photosynthetic stems of cacti. Annals of Botany 70: 317 – 324. Mauseth, J. D. 1993. Medullary bundles and the evolution of cacti. American Journal of Botany 80: 928 – 932.
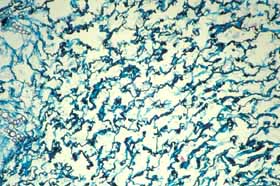
Collapsible water-storage cells. Cactus cortex is divided into an inner water-storing region and an outer photosynthetic layer. More than cortical bundles is needed to move water from the storage region to the chlorophyllous region: water-storing cells should give up water more easily than do other cells. The selective advantage of storing water is not just that it keeps water-storage cells alive but rather that the water can be made available to cells of the photosynthetic palisade cortex, the apical and axillary buds, any flower buds or developing fruits and so on. Consequently, water-storage cells should have thin, flexible walls that can contract or shrink readily such that the cell’s volume diminishes as water is transferred out. On the other hand, the cells that need the water should be more resistant to shrinkage: if all cell walls were equally flexible, all parts of the plant would suffer water stress equally, but that is not adaptive. Instead, water-storage tissues should give up water so easily that the more active cells do not experience water stress unless drought is extremely prolonged. In all cacti, cell walls of the inner cortex are especially thin and flexible, but in many cacti there is an additional modification: the walls are folded or undulate, even when young and recently produced by the shoot apical meristem. Because the walls already have folds in them, the cells are presumably set to shrink very easily. These cells have been called collapsible cortex cells. In contrast, wall of palisade cortex cells – the photosynthetic cells – are thicker and lack folds, so these cannot shrink as easily. Consequently, as the volume of water within a cactus shoot diminishes, cells of the inner cortex give up water preferentially, transferring it to other cells, minimizing water stress and thus allowing photosynthesis to continue.
Mauseth, J. D. 1995a. Collapsible water-storage cells in cacti. Bulletin of the Torrey Botanical Club 122: 145 – 151.
Mucilage cells. A perplexing aspect of water storage in cacti and many other plants is the role of mucilage cells. These are present in many cacti as sparsely dispersed individual cells in some species, as such abundant cells in other species that the plants feel slimy when cut. Spectacular concentrations of mucilage cells occur in prickly pears (members of Opuntia with flat pads) where they form canals many micrometers wide running just exterior to the phloem of the vascular bundles. Plants of living stones (Ariocarpus fissuratus) also have a giant mucilage cavity filling the center of their large taproot. Mucilage is predominately a carbohydrate capable of binding and holding water very firmly. For example, when samples of Opuntia are being prepared for microscopy, they are first dissected, preserved in formaldehyde, then dehydrated by placing them is a series of alcohol solutions, each solution being a stronger solution with less water than the previous one (remember, beer is 5% alcohol, 95% water; wine is 14% alcohol, 86% water). When the formaldehyde is poured off so that the 25% alcohol solution can be poured onto the samples, the mucilage has turned the formaldehyde solution into a slimy mess that – to be really vivid – resembles a runny nose. The mucilage is often so slimy/sticky that the tissue pieces have to be pulled out of it with forceps, it simply will not pour off. This problem continues until the samples have been soaked in 85% alcohol, at which point the slimy/stickiness disappears completely. Even when the samples are exposed to 70% alcohol (140 proof), the mucilage has such a high affinity for water that it can hold the water in the cactus tissue, preventing its loss to the alcohol. You can imagine that the mucilage will be just as effective in a tug-of-war with dry desert air. Undoubtedly, mucilage does help cacti retain water. But with water held tightly in mucilage cells, how does that help all the other cells of the plants? As described with collapsible cortex above, water-storage cells must provide water to the other cells of the plant, not keep it for themselves while the rest of the plant desiccates. One hypothesis is that as cacti experience a prolonged drought, they should be able to digest their mucilage and thus release the water to other cells. If so, the mucilage content of well hydrated plants should differ from that of plants suffering from drought. However, this has not been tested.
Mauseth, J. D. 1980. Release of whole cells of Nopalea (Cactaceae) into secretory canals. Botanical Gazette 141: 15 – 18. Mauseth, J. D. 1980. A stereological morphometric study of the ultrastructure of mucilage cells in Opuntia polyacantha (Cactaceae). Botanical Gazette 141: 373 – 378.
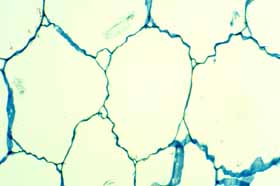
Volume of stem must increase but surface area of epidermis must remain constant. The evolutionary expansion of water-storage tissues in the cortex created problems for the epidermis. Whereas we animals produce new epidermis cells on just about any part of our bodies all through our lives, plants produce new epidermis cells only at shoot and root tips or in developing leaves and flowers.
Consequently, as a succulent stem swells to an increased volume after a rain, it can not increase its surface area correspondingly by producing new epidermis cells. If it were limited by the amount of stretching the epidermis could undergo without splitting open, then the very limited ability to increase surface area would result in a very limited absorption of water. But most cacti – and also many other stem-succulent taxa – have a pleated surface composed of either ribs (vertical pleats only) or tubercles (both vertical and transverse pleats).
It is easiest to think of a cactus cortex as having two parts, an outer cortex consisting of ribs or tubercles and an inner part between the vascular bundles and the bases of the tubercles (see the photo above, right). After the plant has lost water during a prolonged drought, its ribs will have less volume and the two sides of each rib will have pulled together, thus the base of each rib will be narrow. The outer circumference of the inner cortex is the sum of the bases of all the ribs, so with the ribs shrunken, the outer circumference of the inner cortex is smaller, which means that the inner cortex has become thinner through dehydration. When rains return and the plant absorbs water, the inner cortex can swell, becoming thicker and having a larger outer circumference. This increase in outer circumference is accommodated by an expansion of the base of each rib: the volume of both the inner cortex and of the ribs increases whereas the surface area of the shoot remains unchanged. With tubercles instead of ribs, the shoots can cycle between being shorter and taller as well as thinner and thicker. Stem-succulents with pleated surfaces can undergo great changes in volume without any risk of their epidermis being torn open.
Of course, having a pleated surface keeps the shoot from having a minimum S/V ratio, so transpirational surface area is not as low as possible. But the surface area is also important for light absorption, and having a pleated surface increases the amount of light that can be absorbed as well as gives the shoot an ability to contract and expand. As you might guess, the number and height of ribs are important factors: many tall ribs can expand more than a few short ribs. A curious aspect of ribs that is not immediately obvious is that whereas the total surface area of a set of ribs is affected by both the number and height of the ribs, the total volume of water-storage tissues located within the set of ribs is determined only by their height, not by their number. If the number of ribs on a shoot is doubled, then surface area is doubled but the width of the base of each must be reduced to half, so each rib will have half the volume of an original rib. Thus, having numerous ribs of a certain height allows more light absorption (and water loss) than does having fewer ribs of the same height. It might be that stem-succulents of drier habitats would have fewer ribs whereas those of more mesic ones would have more ribs, but this still needs to be examined. Mauseth, J. D. 2000. Theoretical aspects of surface-to-volume ratios and water-storage capacities of succulent shoots. American Journal of Botany 88: 1107 – 1115. Mauseth, J. D. & R. Ross. 1988. Systematic anatomy of the primitive cereoid cactus Leptocereus quadricostatus. Bradleya. 6: 49 – 64. Mauseth, J. D. 1996. Comparative anatomy of tribes Cereeae and Browningieae (Cactaceae). Bradleya 14: 66 – 81. Mauseth, J. D., and J. V. Landrum. 1997. Relictual vegetative anatomical characters in Cactaceae: the genus Pereskia. Journal of Plant Research 110: 55 – 64. Mauseth, J. D., T. Terrazas, and S. Loza-Cornejo. 1998. Anatomy of Relictual Members of Subfamily Cactoideae, IOS Group 1a (Cactaceae). Bradleya 16: 31 – 43.
[End cortex page]
|