| HOME |

|
|
|
Modifications to Cactus
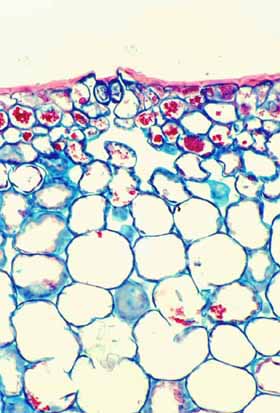
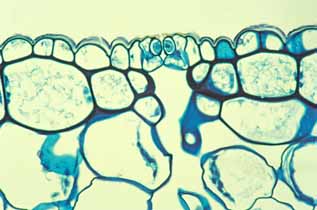
Epidermis Several initial steps in the evolution of cacti to being stem-photosynthetic, stem-succulent plants involved the epidermis. The epidermis is persistent. In most plants, epidermis is an ephemeral tissue that functions for only a few months. This is obviously true of epidermis on leaves that live only from spring until autumn; it is true as well as the stem epidermis of annual plants: no epidermis cell of Arabidopsis thaliana, for example, lives longer than a month and a half or so. And even the stems of perennial plants typically replace their epidermis with bark in either the first or second year of life. But in cacti the epidermis is a long-lived persistent tissue: as long as a cactus stem is green, it still has its epidermis. All cactus bark is tan-color or dark, so it is easy to tell when the shoot loses its epidermis. Giant columnar cacti and huge barrel cacti live for many decades, perhaps even for hundreds of years (plant longevity is an area that needs study), with at most a bit of bark on their lowermost, oldest regions.
Converting epidermis from an ephemeral to a persistent tissue must have involved many substantial changes, certainly more than merely postponing programmed cell death or delaying bark formation. In all cacti, shoot bark is produced by a cork cambium that arises by reactivation of shoot epidermis cells themselves: after decades or centuries of prolonged cell cycle arrest, cactus shoot epidermis cells can be directed to become mitotically active. After prolonged functioning as epidermis cells, they begin to divide with periclinal walls, with the exterior cell of each division differentiating as either a cork cell or a sclereid (most cactus bark consists of alternating thick bands of cork cells and thin bands of sclereids). In a few species, there is even a production of phelloderm to the inner side of the cork cambium. The ability of epidermis cells to function mitotically indicates a variety of remarkable things that could easily be overlooked: their DNA has survived decades of exposure to intense UV irradiation coming through clear desert skies, and all their organelles are healthy and functional. It is also remarkable that epidermis cell protoplasts survive for such a long time while being separated from extraordinarily dry desert air by only a few micrometers of cutinized wall. Another aspect of the evolutionary conversion of epidermis into a persistent tissue almost certainly altered epidermis development. In an ephemeral epidermis, for example that of a leaf or flower petal, the cuticle and waxes produced while the organ is young are sufficient to control water movement for the rest of the life of the organ. But in a long-lived cactus shoot epidermis, the waxes and cuticle produced while the epidermis cells are young and still near the shoot apex probably become weathered away after a year or two and need to be replaced. This is another area that needs research, but it would seem as if cactus shoot epidermis cells must be in a state of “prolonged development,” needing to synthesize cutin and waxes even during the many years they are functionally mature.
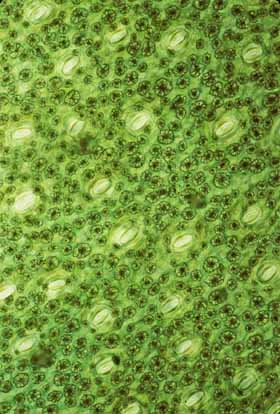
The stem's epidermis has stomata. An obvious feature that the persistent epidermis of a stem-photosynthetic, leafless succulent must have is stomata. Although stem epidermis in many species do have stomata, Urs Eggli in Zurich discovered that stems of the early cacti may have lacked stomata in their epidermis: plants of Pereskia almost completely lack stomata in their stem epidermis (Pereskia is the cactus genus whose members – rather ordinary woody, leafy trees – retain the greatest number of relictual features). Maurizio Sajeva, however, found that in contrast to Pereskia, the stems of all ordinary cacti (that is, the members of subfamilies Cactoideae and Opuntioideae – the cacti that look obviously like cacti) do have stomata and at densities almost as high as in the lower epidermis of Pereskia leaves. If the early evolution of cacti involved obtaining the ability to produce stomata where they had not occurred before, that may have been a type of homeotic evolution: mutations in the promoter regions of stomatal complex morphogenesis genes could have allowed those genes to be activated in stem epidermis as well as in leaf epidermis. Stem epidermis could have more or less instantaneously obtained not only the ability to produce guard cells and subsidiary cells but also the morphogenetic metabolism necessary to control their density and spacing relative to each other.
Mauseth, J. D. 1998. Ontogenetic mechanisms and the evolution of Cactaceae. in: Proceedings of the IV Congreso Latinoamericano de Botánica. Editors: R. Fortunato and N. Bacigalupo. Missouri Botanical Garden Press. pp. 355 – 362. Terrazas Salgado, T, and J. D. Mauseth. 2002. Chapter 2. Shoot anatomy and morphology. in Cacti: biology and uses. (pp 23-40) Edited by P. S. Nobel. University of California Press. Mauseth, J. D. 2004. Cacti and other succulents: stem anatomy of “other succulents” has little in common with that of cacti. Bradleya 22: 131-140. Mauseth, J. D. in press. Blossfeldia lacks cortical bundles and persistent epidermis; is it basal within Cactoideae? Accepted by Bradleya. Mauseth, J. D. 2004. The structure of photosynthetic, succulent stems in plants other than cacti. The International Journal of Plant Sciences 165: 1-9. Mauseth, J. D. 2000. Enfoques anatómicos para el estudio de la biodiversidad: La diversificación de las Cactaceae. In Enfoques contemporáneos para el estudio de la biodiversidad. Eds. H. J Hernández, A. N. García Aldrete, F. Álvarez and M. Ulloa. Instituto de Biología, UNAM, México, México. Mauseth, J. D. 1995. Ontogenetic mechanisms and the evolution of Cactaceae. Giornale Botanico Italiano 129: 429 – 435. [End of Epidermis page]
|