| HOME |

|
|
|
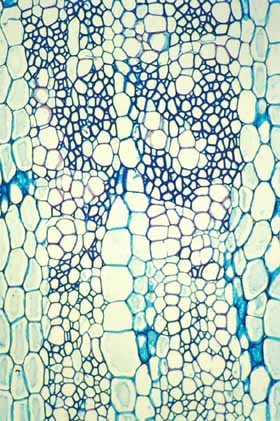
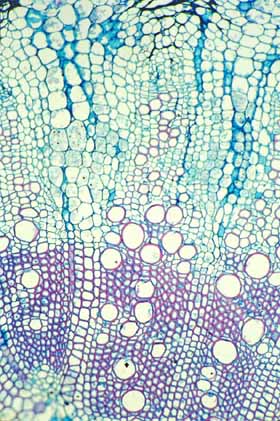
Modifications to Cactus Wood Wood within Cactaceae is amazingly varied. It might be safe to say that no other family has as many types of wood present within it. Ordinary fibrous wood Many cacti have a rather ordinary fibrous wood similar to that in many dicot trees. The wood has a ray system produced by ray initials and an axial system produced by fusiform initials. The rays contain just ray parenchyma cells (although these are often sclerified, having lignified secondary walls) and the axial system is composed of small clusters of vessels, a bit of paratracheal parenchyma and a matrix of xylem fibers. This type of wood occurs in all Pereskia (the leafy trees), and all cacti with such tall, slender bodies that they must have a strong wood to support themselves. The only unusual feature of this wood is that in virtually all examples that have been studied, the fibers are septate and nucleate: they remain alive rather than dying once they have completed their morphogenesis. The rays vary somewhat in height and width and occasionally have cells with rather thick, lignified walls, but the rays are not sufficiently abundant or voluminous to store large amounts of water within the wood. Similarly, the axial parenchyma is limited to just a few cells in immediate contact with vessels (technically known as “scanty paratracheal parenchyma”). The second basic type of axial parenchyma in wood (“apotracheal parenchyma,” surrounded by fibers and not in contact with vessels) does not occur in cacti with fibrous wood.
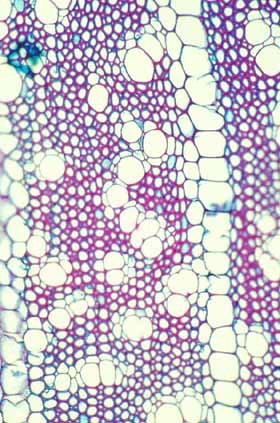
Evolutionary increase in the size of rays An overriding trend in the evolution of cactus wood has been an increase in the amount of water stored within the wood itself. Such water is closer to vessels than is the water stored in the cortex or pith. Increased water-storage capacity in wood has been achieved by different means in different evolutionary lines. In several genera, rays have become gigantic, being several centimeters tall and several millimeters wide: large enough to be easily visible to the naked eye. Assuming that vessels are many centimeters long (perhaps even a meter long), each vessel could have extensive contact with several of these large rays. The water stored within these bodies is present as a three-dimensional reserve with the cortex, pith, rays and paratracheal water all being interconnected and available to vessels.
Evolutionary increase in the amount of axial parenchyma Although some cacti have increased the size or number of their rays, in other species it is mostly the paratracheal parenchyma that has increased in volume. This has occurred to the point where vessels and vessel clusters are now surrounded by abundant paratracheal parenchyma. During times of water stress, vessels do not have to rely on the roots’ ability to extract water from the soil, the vessels can instead quickly and easily pull water from the parenchyma cells that encase them: cavitation should be almost impossible. And if cavitation does occur, the abundant paratracheal parenchyma should be able to refill the vessels. The increase in axial parenchyma is usually accompanied by a decrease in xylem fibers, such that the wood becomes both heavier and weaker; because of this, parenchymatous wood usually occurs in shorter, broader plants in which some of the weight is supported by turgor pressure. Very rarely, the wood matrix consists of just xylem parenchyma with no fibers at all: vessels in plants with such wood traverse nothing but water-storage tissue, all cells are filled with water and the plant would probably die of desiccation before its vessels could possibly cavitate (but this has not been studied).
Alterations in vessel population characters In addition to having water-storage parenchyma near vessels, the wood can be protected from cavitation by altering the ratio of wide vessels to narrow ones. Within any particular plant subjected to water stress, wide vessels tend to cavitate first, narrow ones last. It might seem that a desert-adapted wood should have only very narrow, cavitation-resistant vessels, but narrow vessels have low conducting capacity and have high friction. In contrast, wide vessels can conduct a great deal of water with low friction but they cavitate easily. Which should a plant use? Or should it use a mix? This has been studied in cacti by Josh Stevenson, who developed the concept of the VC50 and VC90; these are the percent of vessels that still are water-filled and functional when the plant has suffered enough drought that cavitation has caused it to lose 50% or 90% of its conducting capacity. With a high VC90, for example, narrow vessels dominate the wood, and as drought begins to be serious, the widest vessels will cavitate first. But because they are so rare, their cavitation does not cause a great loss in water conducting capacity. As the drought continues, narrower and narrower vessels cavitate, but even after 90% of all conducting capacity has been lost, plants with a high VC90 will still have a large number of very narrow vessels left – they are too narrow to conduct much water (but there is not much water available to conduct), and probably that water goes to important areas such as buds and apical meristems (but that has not been tested yet). This appears to be adaptive for cacti. But in plants with a low VC90, wide vessels dominate the wood, and this wood can conduct large amounts of water easily when it is available. But drought will cause the wide vessels to cavitate, and because they are so abundant, the plant will soon have lost use of most vessels -- the only ones that remain water-filled and functional are the rare narrow vessels. The plant may still face months of drought with only a few vessels left; this does not seem optimum for cacti, to put it mildly. Stevenson, J. F., and J. D. Mauseth. 2001. Cavitation and embolism repair in the dimorphic wood of Rhipsalis lumbricoides (Cactaceae). Haseltonia 8: 51 – 57. Stevenson, J. F., and J. D. Mauseth. 2004. Effects of environment on vessel characters in cactus wood. The International Journal of Plant Sciences 165: 347-357. Mauseth, J. D., and J. F. Stevenson. 2004. Theoretical considerations of vessel diameter and conductive safety in populations of vessels. The International Journal of Plant Sciences 165: 359-368.
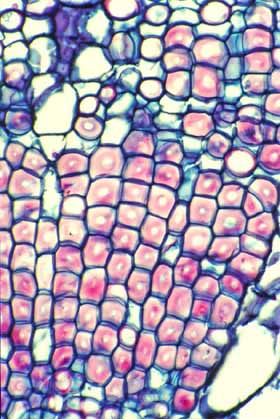
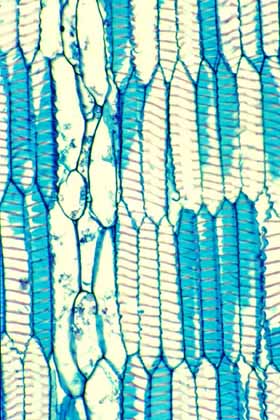
Wide-Band Tracheids (WBTs) An unusual feature in the evolution of cacti is the production of a cell called a wide-band tracheid (WBT). These are short, broad tracheids whose secondary wall has either an annular or a helical pattern but is never scalariform, reticulate or pitted. The “wide-band” in their name comes from the fact that deposition of the secondary wall does not stop once the wall is 1 or 2µm thick but instead continues until the wall almost occludes the lumen. In WBTs with annular secondary walls, it is not unusual to encounter cells with such a narrow lumen remaining that water conduction would be almost impossible.
The main role of WBTs appears instead to be water storage. Because the secondary wall is either annular or helical, most of the primary wall is unlignified and unobstructed so water can enter and leave the cell easily, but more importantly, the rings of the annular type and the helix of the helical type do not hold the WBT to a rigid, fixed length. As the cell loses water, the thin, flexible primary wall can simply bend inward between the rings or gyres of the helix and the cell becomes shorter. No matter how much water the WBT gives up to surrounding tissues, its own volume can shrink to match the volume of water left. Of course, a cell can cavitate only if it contains a volume of water less than its own rigid volume. By shrinking, the cell avoids cavitation. But for dead cells, what is the advantage to avoiding cavitation? By avoiding cavitation, they maintain complete hydrogen-bonding throughout the mass of water. Once rains come again and these plants have water, they also have completely intact, hydrogen-bonded water columns that can be pulled upward by water tension. A nice touch is that these cells then re-expand to their original length. Obviously, WBTs can shrink only if the wood itself can shrink, so WBTs are found almost exclusively in woods that completely lack fibers. The best bet for finding WBT wood in cacti is to find any globose cactus whose weight is supported by turgor pressure: as these plants lose water in a prolonged drought, their WBTs can shrink and avoid cavitation and their entire bodies can shrink because of their tubercles (the pleated surface mentioned on the Cortex page of this web site). Notice the interaction of so many modifications: pleated surface, thick water-storing, weight-supporting cortex vascularized by cortical bundles, wood that is free of fibers but which contains WBTs and abundant parenchyma.
The evolution of WBTs in cacti added another element to wood organization and made possible an especially wide diversity of woods. In some species, wood now consists of narrow tangential bands of vessels and paratracheal parenchyma alternating with thicker bands of pure WBTs. This wood has the appearance of having growth rings with earlywood consisting of vessels and parenchyma, latewood consisting of WBTs. Variations on this theme still have earlywood of vessels and parenchyma but latewood consisting of either a mixture of WBTs and apotracheal parenchyma cells or even WBTs, vessels and parenchyma cells. A problem with this theory is that often a plant that appears to be many years old will have too few “annual rings.” It could certainly be that these are growth rings of early- and latewood but the plants simply do not make a ring each year. It is well-known that desert trees occasionally fail to make growth rings during unusually dry years, but these cacti have so few rings it would appear that they make wood only during exceptionally good years. This needs to be examined under carefully controlled conditions in which the age of the plants is well-known. WBTs occur in all three subfamilies. They are very rare in the Pereskioideae with their hard trunks with fibrous wood, but are almost universally present in Opuntioideae and Cactoideae. In Cactoideae, WBTs have a distribution that would be expected of a tracheid: they occur only in the axial system of the wood (that part produced by fusiform initials), not in the rays (they also occur in the primary xylem of stele bundles and in cortical and medullary bundles). But their distribution in Opuntioideae is often not at all typical of that of a tracheid: they frequently occur in the axial system where they “should” be, but in many opuntioid species they occur in the rays as well – rays being defined as consisting of short cells produced by ray initials and lying opposite the rays in the secondary phloem. In other species they occur ONLY in the rays, not at all in the axial system, and in more than half the species, they occur in the perimedullary zone (the outermost region of pith and innermost regions of the medullary rays). This may seem like merely a puzzling curiosity, but I do not know of any other group of plants that have tracheids or vessels occurring outside the xylem. Mauseth, J. D., Y. Uozumi, B. J. Plemons, and J. V. Landrum. 1995. Structural and systematic study of an unusual tracheid type in cacti. Journal of Plant Research 108: 517 – 526. Mauseth, J. D. 2004. Wide-band tracheids are present in almost all species of Cactaceae. The Journal of Plant Research 117: 69-76.
Phase change in wood of cacti A really amazing feature of the wood of almost all members of Cactoideae is that it undergoes a phase change. This means that for the first part of its life, a stem or root produces one kind of wood, at a later stage of its life it produces a different kind of wood. In almost all (but there are a few exceptions) species of Cactoideae, once a seed germinates and begins to produce wood, it produces WBT wood: it has an axial system of vessels, parenchyma and WBTs but no fibers at all and its rays are very soft parenchyma cells. At some point it undergoes its phase change and begins to make wood in which the axial system is dominated by fibers rather than WBTs. At the same time, vessel characters may change also, and ray cells in the second phase often have thicker walls than those in the first phase. The change can be very abrupt, or it can be gradual with at first the production of a few clusters of fibers near (but usually never touching) WBTs, then more fibers and fewer WBTs until finally no more WBTs are ever produced.
We have no idea what the stimulus is that triggers the change, but it may be related to weight stress. In species whose adults tend to be large and heavy or long and slender, the phase change occurs while the plants are still young and small, less than 10 or 20 cm tall, but which are so slender they tilt and put stress on the wood. Conversely, in species whose seedlings and adults are globose, the broad, squat bodies are always supported by turgor pressure, there is no tilting and there is never any phase change at all, the plant produces only WBT wood its entire life. For species whose adults are of intermediate height, the initial phase of WBT wood is prolonged but not permanent: apparently they produce WBT wood as long as they are short and wide enough to be supported by turgor, but once they finally grow so tall that they begin to lean and must have the skeletal support of strong wood, they switch to making fibrous wood (this is only a hypothesis and has not been tested to verify it). A transverse section at the base of an old plant would reveal WBT wood in the center, fibrous wood near the periphery; a transverse section near the apex would reveal only fibrous wood. One of the themes of this website is the complexity of cactus evolution, and wood dimorphism offers another example. In the genus Samaipaticereus, wood dimorphism runs in the opposite direction: young shoots have hard fibrous wood, older ones have just parenchymatous wood without fibers. So, how is it that an older plant needs less support than a younger one? Samaipaticereus is basically a “support parasite:” it has long slender stems that must be supported by a hard, fibrous wood, but it grows in forests where its long stems soon extend upward into the branches of shrubs and trees. At this point, if it happens to bend too much, it simply ends up leaning against a tree branch and thus no longer needs to support itself, the tree is doing that. At that point, it is advantageous for it to stop expending so much carbohydrate making unnecessary wood fibers, and instead it just makes parenchymatous wood, basically just vessels. This is another point that needs to be documented: we do know for certain that Samaipaticereus has this form of dimorphic wood, but we do not know if it is triggered by sensing that it is leaning against a tree branch or if it just switches to making soft wood and once it falls, it probably falls on something that can hold it up. This is perhaps not truly a case of being a “support parasite” but rather it might be a mutualism: it could be that the tree is protected from browsers by having a mass of spiny Samaipaticereus branches tangled up with its own otherwise edible twigs and leaves.
Mauseth, J. D. 1993. Water-storing and cavitation-preventing adaptations in wood of cacti. Annals of Botany 72: 81 – 89. Mauseth, J. D., and T. Fujii. 1994. Resin-casing: A method for investigating apoplastic spaces. American Journal of Botany 81: 104 – 110. Mauseth, J. D., and B. J. Plemons. 1995. Developmentally variable, polymorphic woods in cacti. American Journal of Botany 82: 1199 – 1205. Mauseth, J. D., and B. J. Plemons-Rodriguez. 1997. Presence of paratracheal water storage tissue does not alter vessel characters in cactus wood. American Journal of Botany 84: 815 – 822. Mauseth, J. D., and B. J. Plemons-Rodriguez. 1998. Evolution of extreme xeromorphic characters in wood: A study of nine evolutionary lines in Cactaceae. American Journal of Botany 85: 209 – 218. Arnold, D. H., and J. D. Mauseth. 1999. Effects of environmental factors on development of wood. American Journal of Botany 86: 367 – 371. Mauseth, J. D. 1999. Anatomical adaptations to xeric conditions in Maihuenia (Cactaceae), a relictual, leaf-bearing cactus. The Journal of Plant Research 112: 307 – 315.
[End Wood page]
|