| HOME |

|
|
|
Modifications to Shoot Apical
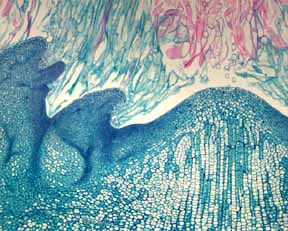
Meristems The evolution of stem-succulence in cacti altered selection pressures acting on shoot apical meristems (SAMs). Think of a piece of vine about three meters long (and a few millimeters wide) compared to a section of shoot of a giant columnar cactus also 3 m long (and maybe 400 mm wide): which plant contains more epidermis cells (just consider the stem epidermis, not the leaf epidermis, which is produced by leaf meristems); which has more cortex cells; which has more pith cells? You can immediately appreciate that, compared to the apex of a vine, the cactus shoot apical meristem will have to produce vastly more cells to construct any particular length of stem. But actually it is worse than that. The succulent stem is much heavier than the narrow stems of most plants, and consequently there has been a great reduction in the amount of branching: if every branch and twig of an ordinary non-succulent tree (such as an oak or a maple for example) were as wide and heavy as a branch of a columnar cactus, the oak or maple would either just fall apart or would crush its trunk. Instead, cacti branch much less than most other plants (palms excepted) and many have just a single stem, and thus just a single shoot apical meristem. That means that whereas the primary body (all the slender twigs and root tips) of a non-succulent tree is constructed from the activity of thousands of shoot apical meristems, the primary body of cacti is produced by just a few, or even by just a single shoot apex.
Having a greatly reduced number of shoot apical meristems increases risk in several ways. Most obvious of course is that damage to one twig out of thousands is negligible whereas damage to one shoot tip out of just a dozen (or just one) is serious. But all the spine clusters of cacti are axillary buds and each has a dormant shoot apical meristem, so damaged meristems can be replaced in cacti as easily as in any other plant. The real threat is to the DNA in the meristem cells. An important concept is not the total number of body cells that must be produced by a meristem but rather the total number of body cells produced by each meristem cell, as discussed by Dr. E. J. Klekowski. Each twig in a non-succulent tree is short and slender and contains just a few epidermis, cortex and pith cells ("just a few" being less than a few hundred or thousand), and the work of producing that small number of cells is divided among several hundred meristem cells. If the whole tree is considered, all those thousands of twigs together must contain close to a million meristem cells. But the evolutionary conversion of narrow stems into giant ones in cacti put added work on the meristem cells: if there were no other change, then each shoot apical meristem cell of a cactus would have to produce vastly more cells than any SAM cell of a maple or other plant. And every time a meristem cell replicates its DNA, there is the chance that a copy-error mutation will be introduced into the meristem cell's nucleus. As the number of cell divisions per cell increases, the chance of damaging the DNA increases. But there was compensatory evolution in the SAMs of cacti: they too increased in size to the point that now some cacti have the largest shoot apical meristems known – 2.0 mm in diameter – large enough to be seen easily with the naked eye. Rather than containing a few hundred meristematic cells as in other plants, cactus apical meristems contain thousands of cells (or maybe tens of thousands – the number has not been calculated yet). This large size brings the number of cell divisions per cell down greatly.
But the evolutionary increase in SAM size had other consequences besides reducing the number of cell divisions per cell: it altered phyllotaxy. Leaf primordia are initiated at the base of shoot apical meristems, so as increased succulence created selective pressure for larger meristems, that resulted in meristems with such large basal circumferences that they have room to produce vastly more leaf primordia than other plants. The size of leaf primordia has not increased as much as has the shoot apical meristem itself, so the increasingly larger basal circumference is occupied by larger numbers of leaf primordia, not by a constant number of larger primordia. Consequently, cacti with giant apical meristems have high phyllotaxy, often very high phyllotaxy, involving numbers high in the Fibonacci series. It is ironic that as cacti were losing the capacity to make large, ordinary leaves, they were gaining the capacity to make huge numbers of leaf primordia.
Molecular genetic studies are revealing the complex interactions between many different genes in the functioning of shoot apical meristems. As cactus meristems evolved to be dramatically larger than those of other plants, especially compared to those of Arabidopsis thaliana, the interaction of those gene products had to be altered as well if the meristems were to continue to function properly. For example, genes such as SHOOT-MERISTEMLESS, WUSCHEL and CLAVATA must control much larger volumes of SAM tissue in cacti than they do in A. thaliana, and genes such as PERIANTHIA might establish much more extensive fields of inhibition around leaf primordia. The giant shoot apical meristems of cacti should make it possible to more accurately map gene expression patterns relative to SAM zonation and leaf primordium sites. Mauseth, J. D. 2004. Giant shoot apical meristems in cacti have ordinary leaf primordia but altered phyllotaxy and shoot diameter. Annals of Botany 94: 145-153. Mauseth, J. D. 1978. An investigation of the morphogenetic mechanisms which control the development of zonation in seedling shoot apical meristems. American Journal of Botany 65: 158 – 167. Mauseth, J. D. 1978. An investigation of the phylogenetic and ontogenetic variability of shoot apical meristems in the Cactaceae. American Journal of Botany 65: 326 – 333. Mauseth, J. D. 1979. Cytokinin‑elicited formation of the pith‑rib meristem and other effects of growth regulators on the morphogenesis of Echinocereus (Cactaceae) seedling shoot apical meristems. American Journal of Botany 66: 446 – 451. Mauseth, J. D., and K. J. Niklas. 1979. Constancy of relative volumes of zones in shoot apical meristems in Cactaceae: implications concerning meristem size, shape, and metabolism. American Journal of Botany 66: 933 – 939. Niklas, K. J., and J. D. Mauseth. 1980. Simulations of cell dimensions in shoot apical meristems: implications concerning zonate apices. American Journal of Botany 67: 715 – 732. Niklas, K. J., and J. D. Mauseth. 1981. Relationships among shoot apical meristem ontogenic features in Trichocereus pachanoi and Melocactus matanzanus (Cactaceae). American Journal of Botany 68: 100 – 105.
[end Apical Meristems page]
|