Cellulose synthesis is one of the most important events in plant growth since it determines both the rate of growth and morphology of the cell. Cellulose synthase is the primary enzyme responsible for synthesis of the biopolymer cellulose (1,4-b-D-glucan). The gene responsible for the coding of cellulose synthase was first found in Acetobacter xylinum and was named celA (Saxena et al, Plant Mol. Biol. 1990 [8]). Homologs of the celA gene were later found in Gossypium hirsutum (cotton) and Oryza sativa (rice) (Pear et al, PNAS. 1996 [9]). A partial length ( 1.3 kb) cDNA was obtained from an A. thaliana library and sequenced. A homology search was performed and high similarity existed between the cotton celA gene sequence and the isolated cDNA (68% identity and 81% similarity). Within the partial cDNA sequence, two of the four highly conserved glycosyltransferase subdomains (U-3 and U-4) were found confirming the possibility that this clone could be a member of the celA gene family. Northern analysis using the 1.3 kb partial cDNA as a probe showed high levels of expression of a 2.2 kb transcript in 0-8 hr old wounded tissue; in addition, lower levels of expression were detected in flowers and correspondingly, whole plants. Genomic Southern blot analysis in A.thaliana using the same probe demonstrated that this cDNA is most likely a single copy, non-multigenic gene.
Introduction
Cellulose synthase is one of the key enzymes in the cellulose biosynthesis pathway and is responsible for forming the 1,4-b-D-glucan bond through its glycosyltransferase activity [13]. Due to its important role in synthesizing the most abundant biopolymer on earth the cellulose synthase gene has been a very sought after target. However, until Saxena et al [8] found a gene responsible for cellulose synthase in Acetobacter xylinum, little was understood about the molecular biology of cellulose synthase in higher plants. Several studies were done to find a motif common to glycosyltransferases (which includes cellulose synthase) and a consensus sequence was found [10]. Further studies were done to introduce a model for the catalytic site using this motif that would be biochemically conceivable [11,12]. It was determined that the catalytic site of celA, which is responsible for binding UDP-Glc, consists of three conserved aspartic acid residues followed by a QXXRW motif. Using this data it was possible for Pear et al [9] to find celA in higher plants (cotton, rice). As expected, all four of the subdomains (U-1,U-2,U-3 and U-4) of celA (containing the necessary aspartic acid residues and QXXRW motif) were highly conserved; however, unlike the Acetobacter cDNA, the cDNAs from the higher plants had regions of hyper variable repeats indicating that conservation of sequence in other regions of the gene was not as crucial.
The celA family of genes have not been well documented in Arabidopsis thaliana up to this point. Finding celA in this model plant system could lead to a better understanding of the characteristics of cellulose synthase and the cellulose biosynthesis pathway in higher plants. This paper attempts to characterize a partial cDNA with high homology to two of the four subdomain regions needed for the celA catalytic site discussed above. Although most of the molecular analysis of the A.thaliana cellulose synthase has yet to be performed this work lays a solid foundation for further experimentation.
Materials and Methods
Isolation and subcloning of cellulose synthesis cDNA:
The cDNA library clone contained in a pZL1 vector (Gibco, Life Technologies) was obtained from the Arabidopsis Biotechnology Research Center. The plasmid containing the cloned cDNA was then isolated from E.coli using a variation on an alkaline extraction procedure [1,2]. Once the plasmid was isolated, the cDNA clone was excised via a restriction digestion using HindIII and EcoRI (Promega). The digestion was checked on an agarose gel and a 1.3 kb fragment was gel eluted using QiagenĽs QIAquick Gel Extraction Kit (Qiagen Inc.) [3]. The isolated 1.3 kb fragment was then subcloned into pBluescript (Stratagene) and the subsequent recombinant vector was transformed into E. coli (DH5a and XL1-Blue) [4]. Transformants were selected using Ampicillin and positive recombinants were „blue/whiteľ screened using X-gal and IPTG. Positives were further checked via miniprep and PCR analysis [5] using T-7 and T-3 primers.
DNA Sequencing and Analysis:
A Sanger dideoxy sequencing reaction [6] was performed on the insert in pBluescript using the Sequenase, version 2.0 DNA Sequencing Kit (United States Biochem.) [7]. T-3 and T-7 were used as primers. In addition automated sequencing of the original pZL1 clone was done at the University of Texas Institute for Cellular and Molecular Biology using SP6 and T-7 as primers. Sequence analysis was done using NCBI's Blast sequence similarity search, Gene Runner, MacVector (Kodak Co.) and SeqApp multiple alignment tool.
RNA isolation from A. thaliana and Northern analysis:
RNA was isolated from whole plants, flowers, stems and leaves. RNA was also isolated from tissue 0,1,5 and 8 hours after wounding. A phenol/chloroform extraction was performed in order to isolate the RNA [15]. A probe was generated and labeled with a-[P32] using the random priming method [16] on the 1.3 kb partial cDNA clone. This cDNA had been amplified directly from a pZL1 vector using PCR. The total RNA was run on 1.2% agarose formaldehyde gel and transferred to a Zetaprobe nylon membrane (BioRad). Hybridization was carried out at 65 degrees C followed by a low stringency wash with 5% SDS, Na2HPO4 and EDTA (also at 65 degrees C). The blot was exposed to film for 4 days.
Isolation of Genomic DNA from A. thaliana and Southern analysis:
Genomic DNA was isolated from whole plant tissue (frozen in liquid N2) using SDS/NaOH and SDS/phenol followed by PEG precipitation [17]. Three separate genomic DNA digestions were performed using EcoRI, HindIII and BamHI and each was run on a 0.8% agarose gel. An alkaline transfer was carried out and the DNA was transferred to a Zetaprobe nylon membrane. This blot was hybridized with the same 1.3 kb probe used for the Northern blot (at 65 degrees C). The low stringency washed used in the Northern was also used on this blot. The blot was exposed to film for 4 days.
Results
Sequencing and analysis:
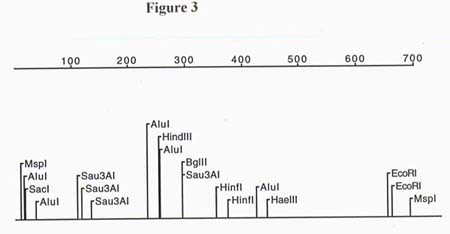
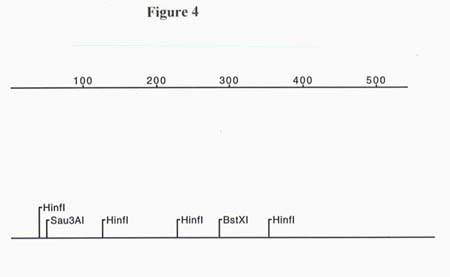
Through restriction digestion of pZL1 it was determined that the partial cDNA clone was 1.3 kb. A total of 1241 bp were sequenced from pZL1 using T-7 and SP6 primers. Due to the limitations on automated sequencing length, the full 1.3 kb insert was not obtained (sequence info. in the middle of the partial length cDNA is missing). Restriction maps of both ends of the cDNA clone were generated and are presented in figures 3 and 4. Since the orientation of the sequence was known due to the poly-A tail, a total of 3 ORFs were analyzed. The only reading frames that made sense for both the 5' and 3' ends resulted in a high score for identity to homologs of cellulose synthase after an NCBI Blast search. The translated ORF for the 5' end of the 1.3 kb partial cDNA started on bp # 2 and had 68% identity and 81% similarity to the Gossypium hirsutum cellulose synthase gene, a 57% identity and 71% similarity to the Agrobacterium tumefaciens cellulose synthase gene and a 52% identity and 68% similarity to Acetobacter xylinum cellulose synthase gene. The BLAST search performed with the ORF translated from the 3' end of the 1.3 kb partial cDNA showed 58% identity and 66% similarity to the celA gene from Gossypium hirsutum. A multiple alignment was done to further examine the homology between the known celA genes from other organisms and the cloned partial cDNA from A. thaliana (see Figure 5). The alignment showed that the highly conserved QXXRW motif , in the U-4 subdomain, of all four organisms matches up with the exclusion of a histidine (H) substituted for a glutamine (Q) on the Arabidopsis thaliana cDNA. This substitution might be explained by an error in sequencing since the codons for histidine are, CAC and CAU, and the codons for glutamine are CAA and CAG. This means that one nucleotide substitution could have resulted in this error. In fact, when the original sequence fluorogram was analyzed there was indeed a C in the third position of the codon but it was a weak signal. In addition to the QXXRW motif there is 100% alignment between all the organisms with regards to one of the three crucial aspartic acid residues located in the U-3 subdomain 38 amino acids 5' of the QXXRW site. Since the 5' end of the cloned cDNA is incomplete due to the priming strategy used when constructing the cDNA library, the remaining two aspartic acid residues (as well as the crucial U-1 and U-2 subdomains) could not be seen or compared.
Southern and Northern analysis:
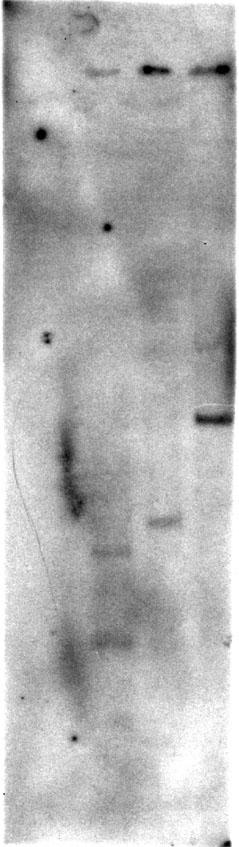
Southern blot analysis (figure 7) demonstrates that the 1.3 kb partial cDNA probe was from a single gene family. Restriction analysis of the probe indicates that only EcoRI should have cut the genomic DNA within the length of the probe. Since only two bands, 0.8 and 1.7 kb, were visible in the EcoRI lane and a 2.0 kb and 3.6 kb band was visible in the BamHI and HindIII lanes respectively, it is evident that the probe is only detecting one gene. It is possible that the probe was bound to other genomic DNA fragments but was removed during the washes; however, this is unlikely due to the fact that the wash was low stringency.
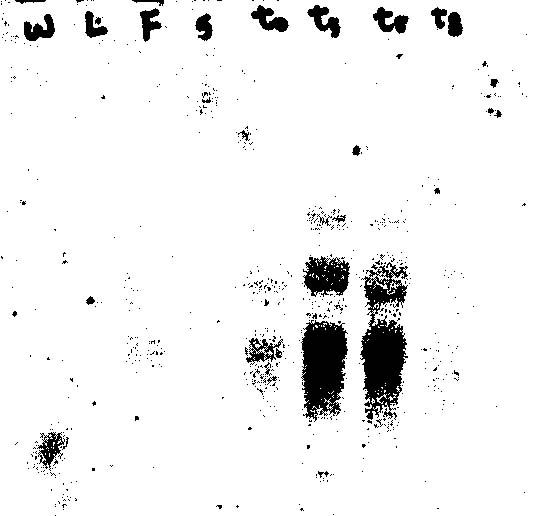
Northern blot analysis (figure 6) indicated high levels of gene expression in tissue that had been harvested up to 5 hours after wounding. Additionally, gene expression was detected in flowers and as a result, a very weak expression signal could be seen from whole plants. The size of the transcript to which the 1.3 kb partial cDNA probe hybridized in both flowers and wounded tissue was approximately 2.2 kb. However, a second band of approximately 4.0 kb was also detected in the RNA from wounded tissue. This band could represent non specific binding to rRNA or a differentially spliced transcript from the same gene that produced the 2.2 kb transcript. Do to the Southern blot analysis which demonstrates that the cDNA probe (hybridized and washed under the same conditions for both blots) is part of a single gene family, it is highly unlikely that the probe was able to detect a totally different but homologous gene product on the Northern.
Discussion
The alignment of the 1.3 kb partial A.thaliana cDNA clone to the G.hirsutum, A.tumefaciens and A.xylinum celA genes provides a strong piece of evidence that suggests the clone is a homolog of the celA genes. Besides having a high alignment to part of the necessary glycosyltransferase UDP-Glc binding motif and the conserved celA subdomains (U-3 and U-4), the 1.3 kb partial cDNA clone shows homology with the less conserved regions of the cotton celA gene such as the 3' end of the HVR region as well as the last 75-100 amino acids. The partial cDNA clone does not show as much homology to the non-conserved regions of A.tumefaciens and A.xylinum but this is to be expected as a result of genetic divergence due to evolutionary distances. One crucial piece of evidence missing in this alignment is the 5' end of the A.thaliana cDNA due to the primers used in the cDNA synthesis step of library construction. This region should contain the crucial U-1 and U-2 conserved subdomains that were not present on the partial cDNA clone. If the 5' end is obtained (via 5' RACE etc.) and found to have high homology to the other known celA genes, then it could be said with much more certainty that this partial cDNA is in fact a member of the celA family.
In G.hirsutum and A.xylinum has been shown that celA is a multigenic gene family [8,9]. The 1.3 kb partial cDNA clone from A.thaliana appears to be a single copy gene as a result of Southern analysis. Since A.thaliana is a higher plant like G.hirsutum, one would expect to find a multigenic celA family in A.thaliana as well. Although this piece of evidence alone does not mean that the partial cDNA discussed here is not a celA gene, it is something to consider.
Northern analysis using the isolated partial cDNA as a probe shows transcript expression in both flowers and wounded tissue. This is consistent with what would be expected from a celA gene. Flowers are actively growing and dividing tissues that would require cellulose biosynthesis. Tissue that has been wounded would also need to cellulose biosynthesis for cells to grow and „healľ the wound. Once wound repair was partially carried out, transcription of celA would diminish. This also correlates with what was seen on the Northern blot. Pear et al [9] have determined that both of the celA gene transcripts in cotton are 3.2 kb long. The Northern analysis of the partial cDNA from A.thaliana shows a transcript that is 2.2 kb long. This could strongly suggest that the 1.3 kb partial cDNA is not in fact a true celA homolog and does not contain crucial conserved regions on the 5' end of the transcript; however, it is also possible that the A.thaliana celA gene doesnĽt have some of the non-conserved regions such as the first 70 amino acids or the P-CR region found in cotton. This issue canĽt be truly resolved until the full A.thaliana cDNA is obtained.
Further work would need to be done to confirm the possibility that this 1.3 kb partial cDNA is indeed a celA transcript. Use of the 5' RACE (rapid amplification of cDNA ends) procedure or other strategies could be utilized to obtain the full length cDNA. Once the full length sequence is obtained and analyzed, an overexpression of the celA cDNA followed by an assay for UDP-Glc binding, could be carried out to back up the sequence analysis.
References
1 Birnboim, H.C. A rapid alkaline extraction method for the isolation of plasmid DNA. Methods. Enzymol. 100:243-255. (1983).
2 Protocol modified by Drs. Paul Krieg and Doug Melton, Harvard University. Protocols and Applications Guide. 2nd Edition, Promega.
3 Qiagen Protocols and Applications Guide. Qiagen Inc., Chatsworth, CA (1995).
4 Instructions on the use of frozen competent cells. Gibco-BRL.
5 Innis, M.A., Gelfand, D.H., Sninsky, J.J., and White, T.J., PCR Protocols. A guide to methods and applications. Academic Press. pp3-13 (1990).
6 Sanger, F.S. Nicklen and A.R. Coulson. DNA Sequencing with chain terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74:5463-5467. (1977).
7 Sequenase Version 2.0 DNA Sequencing Kit. Step by Step Protocols. United States Biochemical, Cleveland, Ohio. pp.1-25.
8 Saxena, I.M.,Lin, F.C. and Brown, R.M., Jr. Plant. Mol. Biol. 15:673-683. (1990).
9 Pear, J.R., Kawagoe, Y., Schreckengost, W.E., Delmer, D.P., and Stalker, D.M. Higher Plants Contain Homologs of the Bacterial celA Genes Encoding the Catalytic Subunit of Cellulose Synthase. Pro. Natl. Acad. Sci. U.S.A. 93, 22:12637-12642 (1996).
10 Delmer, D.P. and Armor, Y. Plant Cell 7:987-1000 (1995).
11 Saxena, I.M., Kudlicka, K., Okuda, K., and Brown, R.M., Jr. (1994). J. Bacteriol. 176:5735-5752.
12 Saxena, I.M., Brown, R.M., Jr. Fevre, M., Geremia, R.A., and Henrissat, B. J. Bacteriology 177:1419-1424. (1995).
13 Tarchevsky, I.A., Marchenko, G.N. Cellulose: Biosynthesis and Structure. Springer-Verlag, Berlin. pp 53-66. (1991).
14 Brown, Malcolm, Jr., Cellulose and Other Natural Polymer Systems. Plenum Press, New York. (1982).
15 Current Protocols in Molecular Biology. pp 4.3.1-4.34
16 Feinberg, A.P. and B. Vogelstein. Analytical Biochemistry. 137:266 (1983).
17 Schuler, M.A. and R.E. Zelinski. Methods in Plant Molecular Biology. pp. 71-73 Academic Press, Inc., San Diego (1989).
Figure Legend
Figure 1. The sequenced and translated (deduced ORF) partial cDNA for Cellulose Synthase.This sequence starts at the 5' end of the cDNA and was obtained by using the T-7 promoter/primer.
Figure 2. More sequenced and translated (deduced ORF) partial cDNA; however, this is the 3' end of the cDNA with the poly-A tail removed. Note: the two sequences do not overlap and do not add up to the full length of the clone; therefore, there is some missing sequence in the middle.
Figure 3. Restriction map of the 5' end of the cDNA clone.
Figure 4. Restriction map of the 3' end of the cDNA clone.
Figure 5. Multiple alignment using SeqApp between Arabidopsis thaliana and: G. hirsutum, A.xylinum and A. tumefaciens.
Figure 6. Northern blot with A. thaliana RNA isolated from (left to right respectively): wholeplant, leaves, flowers, stems, wounded tissue time 0hr, time 1hr, time 5hr, time 8hr.
Figure 7. Southern blot with A.thaliana digested genomic DNA: EcoRI, BamH1 and HindIII respectively.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}